

传染病传播一直是全球公共卫生领域的重要研究方向。经典的SEIR(susceptible-exposed-infected-recovered)模型[1]通过简化假设描述了单一病原体的传播过程,但随着多病毒同时存在并耦合传播现象日益显著,单病毒模型的局限性逐渐凸显。事实上,多种病毒可能同时在同一宿主或群体中出现,不仅共享或竞争传播资源,还会通过削弱或增强宿主免疫力等机制,形成复杂的协同或竞争关系[2-3],可能影响传染病的暴发模式和传播特征。以流感病毒与呼吸道合胞病毒的流行为例,二者在同一季节可能呈现相互促进或抑制的交互模式,影响疾病的暴发时间、传播规模和危害程度[4-5]。Chen等[6]研究表明H1N1和COVID-19的大流行显著扰乱了流感病毒的季节性传播和抗原演化过程。路雅菲等[7]研究显示流感病毒与其他病原体混合或继发感染时,会增加流感患者的重症率和死亡率。Huang等[8]通过研究患者的临床表现症状,探讨了流感B型病毒和SARS-CoV-2(新冠病毒)的共感染情况。因此,多病毒耦合现象为传染病传播规律和公共卫生风险的评估带来了新的挑战。
传统的研究大多聚焦于单病毒传播[9],较少涉及多病毒耦合传播的复杂机制以及其对疫情传播的影响。为弥补单病毒模型的不足,部分学者尝试引入多病毒耦合传播框架,分析病毒在不同网络或宿主条件下的协同或竞争关系,然而,这些多病毒研究仍存在不足。部分研究未能充分考虑不同病毒株和宿主免疫反应的多样性(如免疫逃逸等)对共感染结果的影响,限制了研究结果的广泛适用性[2-3, 5, 10]。另外,现有多病毒模型更多地聚焦于病毒动力学传播理论推导或数值模拟,对人群行为变化、医疗资源分配等干预措施对多病毒的复杂影响尚缺乏综合性考虑[7, 11]。同时,多病毒模型的验证大多停留在局部实验或模拟环境,对实证数据验证仍显不足[8, 12-13]。
因此,本文提出了一种多病毒耦合传播模型,通过系统分析病毒间的耦合作用,结合宿主免疫反应和环境干预因素等,综合探讨多病毒耦合传播的动力学特征和机制。引入了相互促进与相互抑制、免疫逃逸等多样性因素,提升模型对真实场景的适应性;整合了医疗资源、人员行为变化及防控干预对疫情传播的影响,构建多因素耦合框架;通过模拟实验研究分析病毒间相互作用对病毒传播的影响,并通过实证数据验证模型的有效性,为多病毒传播风险评估和精准防控策略提供科学依据。
1 病毒间相互作用
1.1 相互促进作用
1.2 相互抑制作用
1.3 免疫逃逸作用
2 多病毒耦合传播模型
经典SEIR模型通过将人群划分为4个状态:易感者(S)、暴露者(E)、感染者(I)和恢复者(R),有效地描述了单一病毒的传播动态。然而,随着传染病研究的深入,尤其是在多病毒流行和复杂传播机制下,SEIR模型的局限性逐渐显现。为了更准确地模拟传染病传播过程,特别是在涉及无症状感染和隔离措施的情况下,本文对SEIR模型进行扩展与改进,提出了SEAIDRM(susceptible-exposed-asymptomatic-infected-divided-recovered-mortal)模型,以更准确地表征单一病毒传播过程中复杂的人群状态转移。在此基础上,进一步构建了多病毒耦合传播模型,重点分析了病毒间的相互作用(包括相互促进、相互抑制)对传播动力学的影响。
2.1 SEAIDRM模型
SEAIDRM模型进一步细化了人员状态,增加了无症状感染者(asymptomatic,A)、隔离者(divided,D)、死亡者(mortal,M)3种状态,同时在该模型中I代表有症状感染者。在每个时间步,具有感染性的人(A及未被隔离的I)与S接触后,S将有一定概率β被感染变成E;E可能以概率σA转变成A,或以概率σI转变成I;A进一步可能以概率γA变成R,或以概率θA发展成为I;I可能有一定概率ωI被隔离转变成D;未被隔离的I可能以概率γI变成R;D通过治疗可能以概率γD转变成R,也可能以概率μD变成M。SEAIDRM模型能够更全面地描述病毒传播动态,为研究多病毒耦合传播提供了坚实的模型基础。SEAIDRM模型的人员状态转换如图 1所示。
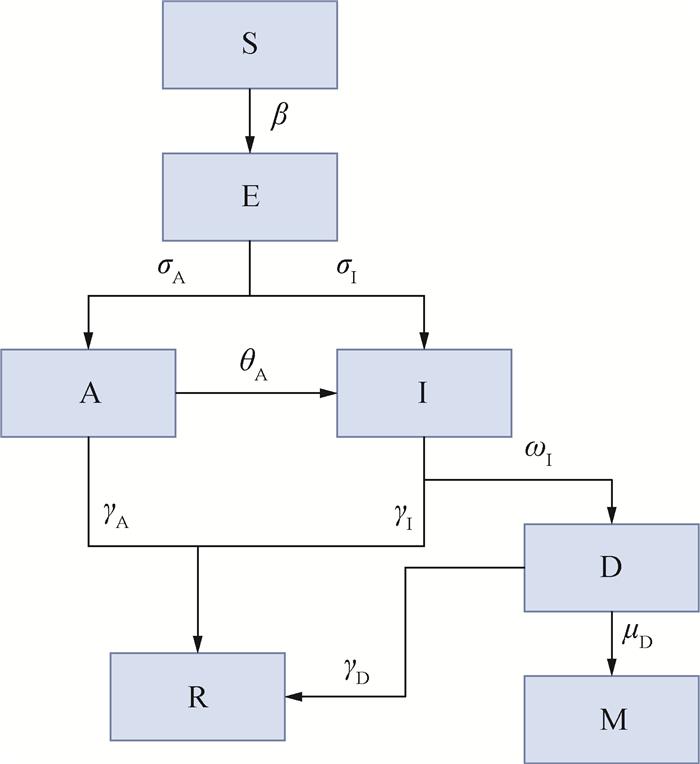
针对给定研究单元,假设总人口不变,描述人群在不同状态间变化的微分方程组为:
其中:S(t)、E(t)、A(t)、I(t)、D(t)、R(t)、M(t)分别代表S、E、A、I、D、R、M在时刻t的数量;t为时间变量;N为人口总数。
2.2 多病毒耦合传播模型
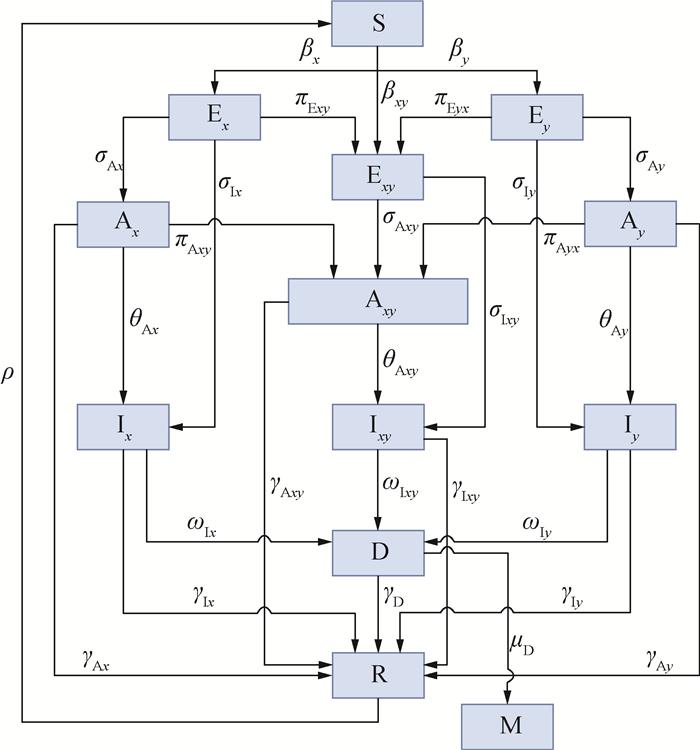
基于式(1)-(7),融合x和y的耦合传播关系,得到x和y耦合传播的常微分方程组为:
x和y耦合传播模型的各参数含义说明如表 1所示。
表 1 多病毒耦合动力学模型参数表 |
| 参数 | 含义 |
| βx | S感染x变成Ex的概率 |
| βy | S感染y变成Ey的概率 |
| βxy | S感染x、y变成Exy的概率 |
| σAx | Ex转变为Ax的概率 |
| σIx | Ex转变为Ix的概率 |
| πExy | Ex转变为Exy的概率 |
| σAy | Ey转变为Ay的概率 |
| σIy | Ey转变为Iy的概率 |
| πEyx | Ey转变为Exy的概率 |
| σAxy | Exy转变为Axy的概率 |
| σIxy | Exy转变为Ixy的概率 |
| γAx | Ax转变为R的概率 |
| θAx | Ax转变为Ix的概率 |
| πAxy | Ax转变为Axy的概率 |
| γAy | Ay转变为R的概率 |
| θAy | Ay转变为Iy的概率 |
| πAyx | Ay转变为Axy的概率 |
| γAxy | Axy转变为R的概率 |
| θAxy | Axy转变为Ixy的概率 |
| γIx | Ix转变为R的概率 |
| ωIx | Ix转变为D的概率 |
| γIy | Iy转变为R的概率 |
| ωIy | Iy转变为D的概率 |
| γIxy | Ixy转变为R的概率 |
| ωIxy | Ixy转变为D的概率 |
| γD | D转变为R的概率 |
| μD | D转变为M的概率 |
| ρ | R再次转变为S的概率 |
2.3 病毒间相互作用对耦合模型的影响
1) 相互促进作用对模型参数的影响。
如果宿主已经感染y,并且y对x有促进作用,则宿主对x的易感性将增强,导致βx增大。
其中:β′x是y促进作用下的S感染x的概率,αyx是y对x传播的促进作用系数,π′Eyx是y促进作用下的Ey接触感染x病毒的人转变为Exy的概率。
同理,如果宿主已经感染x,并且对y有促进作用,则宿主对y的易感性将增强,导致βy增大。
其中:β′y是x促进作用下的S感染y的概率,αxy是x对y传播的促进作用系数,π′Exy是x促进作用下的Ex接触感染y病毒的人转变为Exy的概率。
宿主同时感染2种病毒的概率也显著增大。
其中β′xy是x、y共同促进作用下的S同时感染x、y的概率。
2) 相互抑制作用对模型参数的影响。
如果宿主已经感染y,并且对x有抑制作用,则宿主对x的易感性将降低,导致βx减小。
其中:β″x是x抑制作用下的S感染x的概率,δyx是y对x传播的抑制作用系数,π″Eyx是y抑制作用下的Ey接触感染x的人转变为Exy的概率。
同理,如果宿主已经感染x,并且对y有抑制作用,则宿主对y的易感性将降低,导致βy减小。
其中:β″y是x抑制作用下的S感染y的概率,δxy是x对y传播的抑制作用系数,π″Exy是x抑制作用下的Ex接触感染y的人转变为Exy的概率。
宿主同时感染2种病毒的概率也显著减小。
其中β″xy是x、y共同抑制作用下的S同时感染x、y的概率。
3) 免疫逃逸作用对模型参数的影响。
该模型通过免疫逃逸率ρ量化病毒免疫逃逸机制,其强度受基础逃逸率、免疫人群规模等影响。这种逃逸作用主要体现在使部分恢复人群重新易感、导致免疫力损失等方面,模型利用ρ综合刻画病毒免疫逃逸对疫情演变与传播的影响。
3 模拟实验与结果分析
本节设计了4种场景,来模拟分析相互作用强度、免疫逃逸强度、传播率对多病毒耦合传播的影响。综合不同的病毒性呼吸道传染病(包括新冠、流感、SARS等)的感染特性,包括基本再生数、传染期、潜伏期、治愈时间等,采用的耦合传播模型关键参数如表 2所示。
表 2 x和y耦合传播模型关键参数 |
| 参数 | 取值 | 依据说明 |
| βx | 0.25 | 病毒性呼吸道传染病的基本再生数为1.3~5.0,传染期为3~21 d,βx=基本再生数/(传染期/d),取0.25 |
| βy | 0.18 | 病毒性呼吸道传染病的基本再生数为1.3~5.0,传染期为3~21 d,βy=基本再生数/(传染期/d),取0.18 |
| σAx | 0.20 | 病毒性呼吸道传染病的潜伏期约为1~14 d,σAx=1/(潜伏期/d),取0.20 |
| σIx | 0.10 | 病毒性呼吸道传染病的潜伏期约为1~14 d,σIy=1 /(潜伏期/d),取0.10 |
| σAy | 0.12 | 病毒性呼吸道传染病的潜伏期约为1~14 d,σAy=1/(潜伏期/d),取0.12 |
| σIy | 0.08 | 病毒性呼吸道传染病的潜伏期约为1~14 d,σIy=1/(潜伏期/d),取0.08 |
| γAx | 0.08 | 病毒性呼吸道传染病的无症状感染者平均自愈时间约为3~14 d,γAx=1/(无症状感染者平均自愈时间/d),取0.08 |
| θAx | 0.15 | 病毒性呼吸道传染病的无症状感染者发展为有症状感染者的概率取10%~20% |
| γAy | 0.12 | 病毒性呼吸道传染病的无症状感染者平均自愈时间约为3~14 d,γAy=1/(无症状感染者平均自愈时间/d),取0.12 |
| θAy | 0.10 | 病毒性呼吸道传染病的无症状感染者发展为有症状感染者的概率取10%~20% |
| γIx | 0.07 | 病毒性呼吸道传染病的有症状感染者平均自愈时间约为3~21 d,γIx=1/(有症状感染者平均自愈时间/d),取0.07 |
| ωIx | 0.05 | 病毒性呼吸道传染病的隔离率取5% |
| γIy | 0.08 | 病毒性呼吸道传染病的有症状感染者平均自愈时间约为3~21 d,γIy=1/(有症状感染者平均自愈时间/d),取0.08 |
| ωIy | 0.05 | 病毒性呼吸道传染病的隔离率取5% |
| γD | 0.07 | 隔离后平均恢复时间约为10~21 d,γD=1 / (隔离后平均恢复时间/d),取0.07 |
3.1 模拟场景设置
1) 相互促进作用场景:假设x和y之间存在相互促进作用,能够增强彼此的传播能力。基于式(21)-(25),通过调整相互促进作用强度来模拟不同强度下的传播趋势,分析相互促进对传播规模和速度的影响。
假设x对y传播的促进系数与y对x传播的促进系数保持一致,即αxy=αyx=α,相互促进系数α变化范围为0.1~0.3,越大则促进作用强度越高。
2) 相互抑制作用场景:假设x和y之间存在相互抑制效应,能够减少彼此的传播能力。基于式(26)-(30),通过调整相互抑制作用强度来模拟不同抑制强度下的传播动态,分析相互抑制对疫情传播高峰和整体规模的影响。
假设x对y传播的抑制系数与y对x传播的抑制系数保持一致,即δxy=δyx=δ,相互抑制系数δ变化范围为0.1~0.3,越大则抑制作用强度越高。
3) 免疫逃逸作用场景:假设x和y因基因突变等原因具有免疫逃逸效应,能够使部分恢复人群重新易感、导致免疫力损失等。通过调整免疫逃逸率来模拟不同免疫逃逸强度下的传播动态,分析免疫逃逸作用对疫情传播的影响。
4) 传播率影响作用场景:假设x和y具有不同的传播率,对病毒的传播速度和感染规模具有不同的影响。通过调整βx和βy来模拟不同传播率下的传播动态,分析传播率对疫情传播的影响。
3.2 相互促进强度对多病毒耦合传播影响模拟
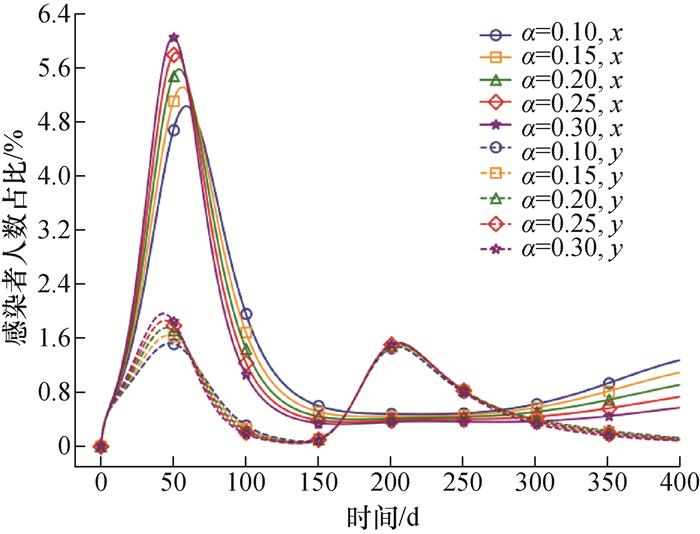
图 3展示了不同α下,x和y感染者人数占比的变化曲线。随着α从0.10增加到0.30,2种病毒的感染高峰均显著提高,且α增大对x的传播增强效果略高于对y的。同时,2种病毒的第一感染高峰均在第50天左右出现,变化趋势相对一致,但x的感染者人数占比的峰值在同一α下略高于y的。此外,2种病毒的感染者人数占比曲线均表现出明显的双峰特性,x的感染者人数占比的第二感染高峰到达时间比y的晚。当x的感染者人数占比在下降的时候,y的感染者人数占比上升,呈现第二高峰,当y的感染者人数占比到达第二高峰后开始下降的时候,x的感染者人数占比开始呈现上升趋势,说明x和y的感染者人数占比在第一个峰值后呈现出交替的波动模式。整体来看,相互促进作用提升了x和y的感染规模,拉长了传播周期,并对x的传播增强效应更为突出。
3.3 相互抑制强度对多病毒耦合传播影响模拟
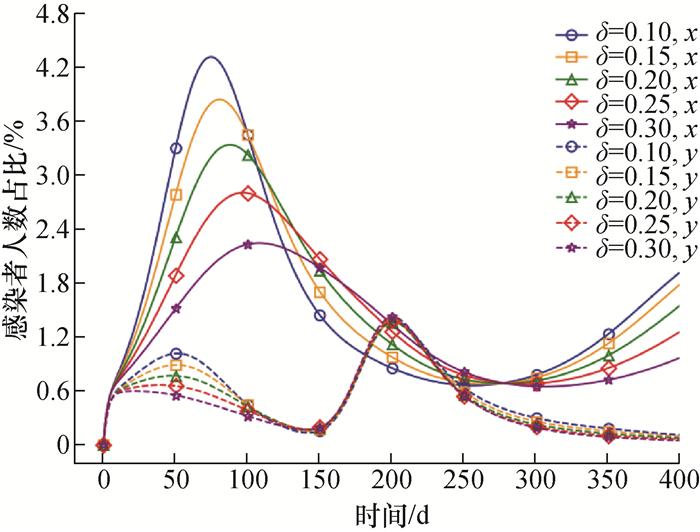
图 4展示了不同δ下,x和y感染者人数占比的变化曲线。随着δ从0.10增加到0.30,2种病毒的感染高峰降低,且δ的增加对x的传播抑制效果明显高于对y的。2种病毒的感染者人数占比曲线均表现出明显的双峰特性,在第一个感染高峰的时候,x和y的感染者人数占比变化趋势相对一致,但y的感染者人数占比的峰值时间相对较早,大约在第50个时间步,x的感染者人数占比的峰值时间约在第70个时间步,且x的感染者人数占比的第一个波峰持续时间较长。在第一个感染高峰后,x的感染者人数占比一直下降,在第270个时间步开始回升,但y的感染者人数占比在第150个时间步开始回升,x和y的感染者人数占比在第一个峰值后同样呈现出交替波动的模式。此外,随着δ的增加,x的感染者人数占比的峰值到达时间逐渐延迟,但y的感染者人数占比的峰值到达时间变化较小。整体来看,相互抑制作用降低了x和y的感染规模,延缓了传播速度,且对x的传播减弱效应较明显。
3.4 免疫逃逸作用对多病毒耦合传播影响模拟
图 5展示了不同ρ下,感染者人数占比(包括只感染x、只感染y及耦合感染x与y)随时间变化的情况。当ρ从0.01增大到0.2时,感染者人数占比显著上升,表明免疫逃逸率的增加能够增强病毒的传播能力。在低免疫逃逸率(ρ=0.01)下,感染者人数占比达到峰值后下降,并维持在较低水平;随着ρ的增大,感染者人数占比的峰值增大,且到达峰值后下降趋势逐渐变缓,呈现出较高且相对平稳的感染者人数占比。在ρ=0.2时,感染者人数占比稳定在9%左右,未出现明显下降。说明免疫逃逸率的提升不仅延长了病毒的传播周期,还增大了长期感染的规模。
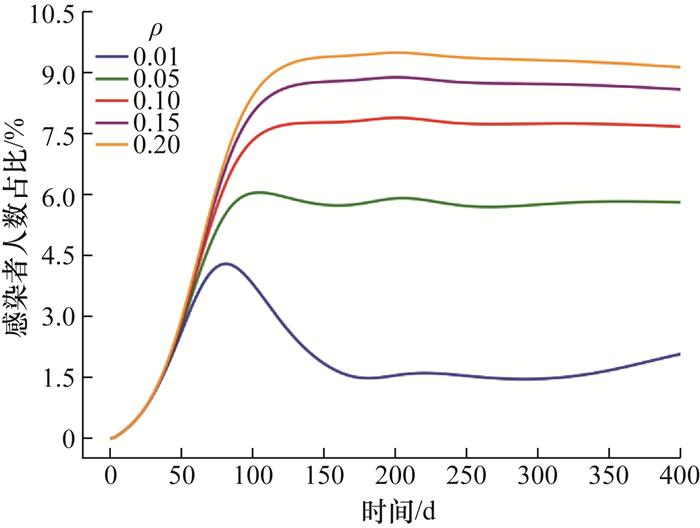
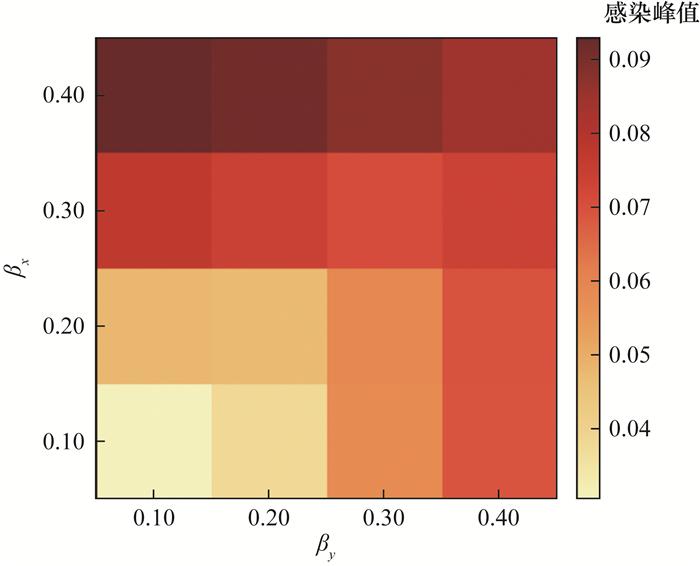
3.5 传播率参数对感染峰值的影响分析
4 实例验证
4.1 数据说明
表 3 SARS-CoV-2和流感病毒耦合传播模型关键参数 |
| 参数 | 取值 | 依据说明 |
| βx | 0.35 | SARS-CoV-2的基本再生数为2.5,传染期为7 d,βx=基本再生数/(传染期/d),取0.35 |
| βy | 0.20 | 流感病毒的基本再生数为1.4,传染期为7 d,βy=基本再生数/(传染期/d),取0.20 |
| σAx | 0.20 | SARS-CoV-2潜伏期约为1~14 d,σAx=1/(潜伏期/d),取0.20 |
| σIx | 0.14 | SARS-CoV-2潜伏期约为1~14 d,σIx= 1/(潜伏期/d),取0.14 |
| σAy | 0.30 | 流感病毒潜伏期约为1~4 d,σAy=1/(潜伏期/d),取0.30 |
| σIy | 0.25 | 流感病毒潜伏期约为1~4 d,σIy=1/(潜伏期/d),取0.25 |
| γAx | 0.07 | SARS-CoV-2无症状感染者平均自愈时间约为7~14 d,γAx=1/(无症状感染者平均自愈时间/d),取0.07 |
| θAx | 0.15 | SARS-CoV-2无症状感染者发展为有症状感染者的概率取10%~20% |
| γAy | 0.20 | 流感病毒无症状感染者平均自愈时间约为3~5 d,γAy=1/(无症状感染者平均自愈时间/d),取0.20 |
| θAy | 0.15 | 流感病毒无症状感染者发展为有症状感染者的概率取10%~20% |
| γIx | 0.05 | SARS-CoV-2有症状感染者平均自愈时间约为14~21 d,γIx=1/(有症状感染者平均自愈时间/d),取0.05 |
| ωIx | 0.10 | SARS-CoV-2隔离率取10% |
| γIy | 0.14 | 流感病毒有症状感染者平均自愈时间约为3~7 d,γIy=1/(有症状感染者平均自愈时间/d),取0.14 |
| ωIy | 0.02 | 流感病毒隔离率取2% |
| γD | 0.07 | 隔离后平均恢复时间约为15 d,γD=1/(隔离后平均恢复时间/d),取0.07 |
备注:x代表SARS-CoV-2;y代表流感病毒。 |
4.2 模拟结果与真实数据对比分析
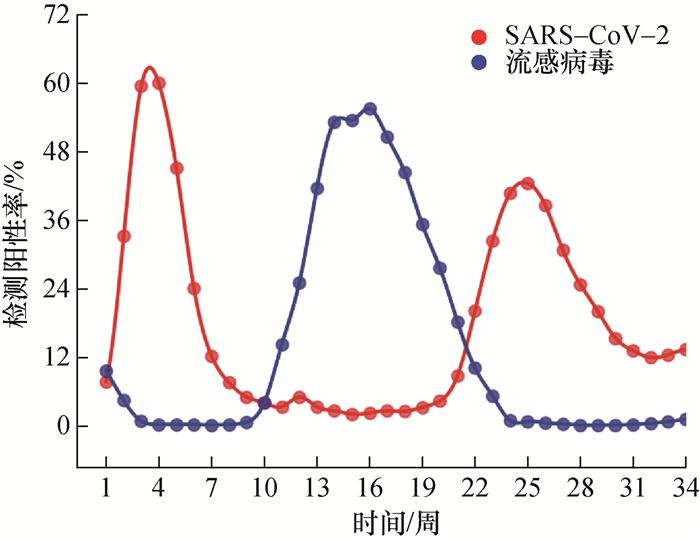
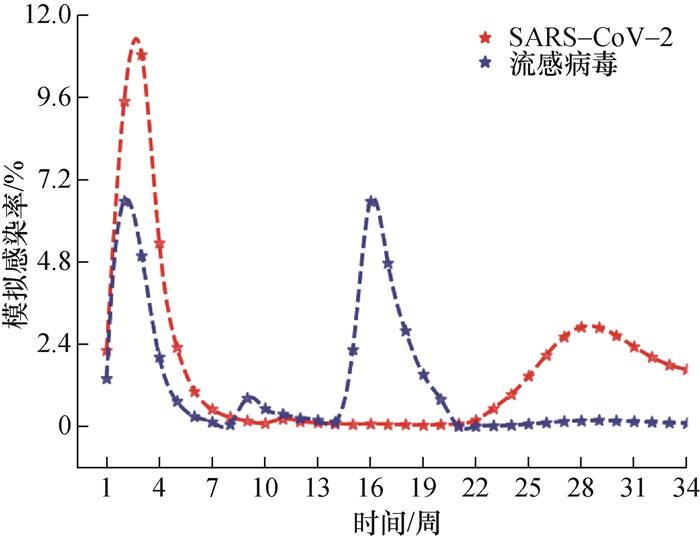
由图 7和8可以看出,SARS-CoV-2的模拟结果在变化趋势上基本与真实数据吻合,呈现双峰的现象;流感病毒的模拟结果与真实数据不同的是在第1~4周也呈现波峰现象,相同的是在第14~20周左右感染情况出现回升。在感染比例上,模拟感染率比检测阳性率低,主要原因可能包括:模型的参数估计可能存在误差;真实数据源自全国哨点医院监测数据,检测阳性率代表病毒样例占门(急)诊就诊人数比例,反映的是就诊患者中检测结果为SARS-CoV-2和流感病毒的平均水平,而模拟感染率代表感染者人数占总人数比例,反映的是病毒在整个研究区域内的总体传播水平。整体来看,耦合传播模型的模拟结果在宏观变化趋势、峰值时间、波峰持续时间等关键指标方面与真实数据相对一致。
4.3 综合干预强度分析
图 9表明不同时间段内综合干预强度是波动变化的。在流行初期(第1~3周),综合干预强度迅速提高,体现了疫情迅猛传播时,需要快速进行防控干预,以应对疫情传播态势。随着感染人数的减少(第7~12周),综合干预强度逐渐下降。在新一轮疫情传播的初期(第13~20周),综合干预强度再次提升,反映了疫情反弹时,需要动态调整防控措施,以防止疫情进一步传播。从曲线的多次下降可以看出,综合干预强度的波动与疫情发展趋势密切相关。
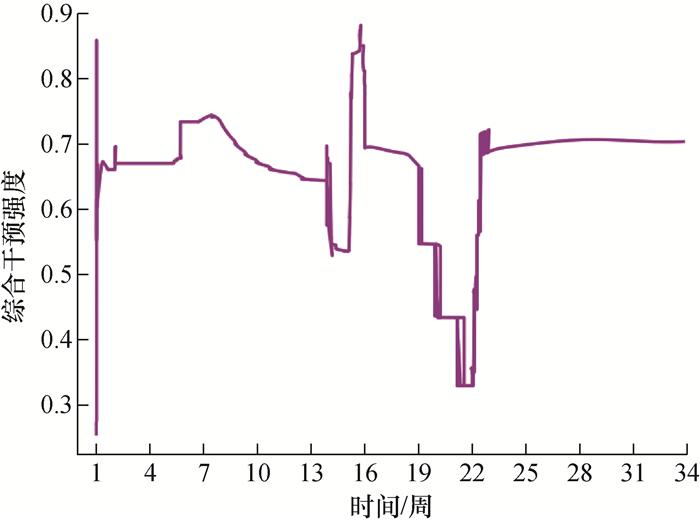
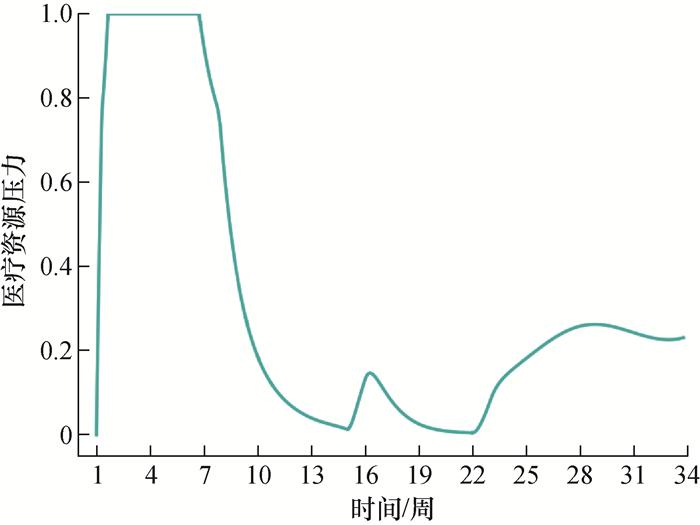
4.4 医疗资源压力分析
5 结论
本文通过对传统SEIR模型的拓展,提出了一种全新的多病毒耦合传播模型SEAIDRM,纳入了无症状感染者、隔离者和死亡者等状态变量,更全面地反映了多病毒共存下的复杂传播动态。该模型有效刻画了病毒间的相互作用与免疫逃逸作用对传染病传播的影响,为传染病耦合传播的研究提供了新的理论视角。
研究结果显示,SEAIDRM模型更符合多病毒共存环境下社会人群的疾病状态变化情况,且模拟结果较好。同时,通过引入相互促进作用系数、相互抑制作用系数和免疫逃逸率,模拟了病毒间的相互作用对传染病耦合传播的影响,发现病毒间的相互促进作用能够提升感染规模,拉长传播周期,导致感染峰值到达的时间提前,同时对感染规模大的病毒的传播增强效应比对感染规模小的病毒的更明显;相互抑制作用可以降低感染规模,延缓传播速度,导致感染峰值到达的时间延后,同时对感染规模大的病毒的传播减弱效应比对感染规模小的病毒的也更明显;免疫逃逸率的提升可以显著增加感染规模,延长病毒的传播周期。研究结果较好地解释了多病毒共存下社会人群的感染规律,强调了不同病毒间复杂的耦合机制在实际传染病预测与分析中的关键性。
本文为多病毒耦合环境中传染病的传播提供了一个新的理论框架,并为流行病风险分析和干预策略研究提供了更准确和适用的模型支持。基于本文的研究结果,可以为公共卫生干预策略的制定提供重要的科学依据。在多病毒共存并耦合传播的情况下,可以通过病毒间的相互作用,制定精细化的防控措施,优化医疗资源配置,以更有效地控制传染病的传播。需要说明的是,本文通过模拟分析发现,x与y耦合传播时,βx的取值可能改变βy与感染峰值的正负相关性,其内在机制与原因需结合动力学模型和实验数据进一步探究,从而为制定精准防控策略提供更有效的依据。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}